G protein


G proteini (guanin nukleotid vezujući proteini) su familija proteina kaja učestvuje u prenosu hemijskih signala izvan ćelije, i uzrokuje promene unutar ćelije. Oni komuniciraju signale mnogih hormona, neurotransmitera, i drugih signalnih faktora.[1]
G protein spregnuti receptori su transmembranski receptori. Signalni molekuli se vezuju za domen lociran izvan ćelije. Intracelularni domen aktivira G proteine. G proteini aktiviraju kaskadu drugih jedinjenja, i što dovodi do ćelijskih promena.
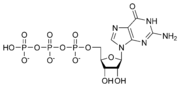
G protein kompleksi se vezuju za fosfatne grupe. Oni funkcionišu kao molekularni prekidači. Kad su vezani za kompleks sa tri fosfatne grupe (guanozin trifosfat (GTP)), oni su “uključeni“, do kad su vezani za kompleks sa samo dve fosfatne grupe (guanozin difosfat (GDP)), oni su isključeni.
G proteini regulišu metaboličke enzime, jonske kanale, transportere, i druge delove ćelijske mašinerije, kontrolišući time transkripciju, motilnost, kontraktilnost, i sekreciju, i time u širem smislu regulišu sistemske funkcije poput embrionskog razvoja, učenja i memorije, i homeostaze.[2]
G proteine su otkrili Alfred G. Gilman i Martin Rodbel tokom njihovog istraživanja ćelijske stimulacije adrenalinom. Oni su utvrdili da kad se adrenalin veže za receptor, receptor ne stimuliše enzime direktno. Umesto toga, stimulacija se odvija kroz G protein, koji stimuliše jedan od enzima, na primer adenilat ciklazu, koja proizvodi sekundarni glasnik ciklični AMP.[3] Za to otkriće, njima je dodeljena Nobelova nagrada za fiziologiju ili medicinu 1994 godine.[4]
G proteini pripadaju većoj grupi enzima zvanih GTPaze.
Funkcija
G proteini su važni signal transdukujući molekuli u ćelijama. Malfunkcija GPCR signalnih puteva dovodi do mnogih bolesti, kao što su dijabetes, slepilo, alergije, depresija, kardiovaskularni defekti, i neke forme raka. Procenjuje se da su GPCR receptori celularne mete za više od polovine modernih lekova.[5]
Ljudski genom sadrži oko 950 G protein-spregnutih receptora, koji detektuju fotone (svetlost), hormone, faktore rasta, lekove, i druge endogene ligande. Približno 150 GPCR receptora nađenih u ljudskom genomu ima nepoznate funkcije.
Tipovi G protein signalizacije
Termin G protein se može odnositi na dve distinktne familije proteina. Heterotrimerni G proteini se ponekad nazivaju velikim G proteinima. Oni su aktivirani G protein-spregnutim receptorima, i sastoje se od alfa (α), beta (β), i gama (γ) podjedinica. Postoje takođe i mali G proteini (20-25 kDa), koji pripadaju Ras superfamiliji malih GTPaza. Ti proteini su homologni sa alfa (α) podjedinicom nađenom kod heterotrimera, ali su monomerni. Međutim, oni takođe vezuju GTP i GDP, i učestvuju u transdukciji signala.
Heterotrimerni G proteini
Razni tipovi heterotrimernih G proteina imaju zajednički mehanizam. Oni se aktiviraju u responsu na konformacionu promenu G protein-spregnutog receptora, razmenjuju GDP za GTP, i disociraju se da bi aktivirali druge proteine u putu prenosa signala. Specifični mehanizmi, međutim, se razlikuju među vrstama.
Zajednički mehanizam

Receptorom-aktivirani G proteini se vezuju za unutrašnju stranu ćelijske membrane. Oni se sastoje od Gα i čvrsto povezane Gβγ podjedinice. Postoji više klasa Gα podjedinica: Gsα (G stimulatori), Giα (G inhibitori), Goα (G drugi), Gq/11α, i G12/13α su neki od njih. Oni se različito ponašaju nakon prepoznavanja efektora, ali imaju sličan mehanizam aktivacije.
Aktivacija
Kad ligand aktivira G protein-spregnuti receptor, on indukuje konformacionu promenu receptora koja dozvoljava receptoru da funkcioniše kao faktor razmene guanin nukleotida (GEF) koji razmenjuje GDP za GTP na Gα podjedinici. Po tradicionalnom shvatanju aktivacije heterotrimernih proteina, ta razmena otpočinje disocijaciju Gα podjedinice, vezane za GTP, od Gβγ dimera i od receptora. Postoje i alternativni modeli koji sugestiraju molekularno preuređivanje, reorganizaciju, i pri-formiranje kompleksa molekulskih efektora.[6][7] Gα-GTP i Gβγ mogu da aktiviraju različite signalne kaskade (ili puteve sekundarnih glasnika) i efektorskih proteina, dok je receptor spreman da aktivira sledeći G protein.
Terminacija
Gα podjedinica će vremenom hidrolizirati vezani GTP u GDP putem svoje inherentne enzimske aktivnosti, čime se omogućava ponovno vezivanje za Gβγ i početak novog ciklusa. RBM (ili RGS) grupa proteina dejstvuje kao GTPaza-aktivirajući proteini (GAP), specifično za Gα podjedinice. Efekat tih proteina je ubrzanje GTP hidrolize u GDP i terminacija prenosa signala. U nekim slučajevima, sam efektor može posedovati unutrašnju GAP aktivnost, što pomaže u deaktiviranju puta. To je slučaj sa fosfolipazom C beta, koja ima GAP aktivnost unutar svog C-terminalnog regiona. To je alternativna forma regulacije Gα podjedinice.
Specifični mehanizmi
- Gαs aktivira cAMP-zavisni put koji stimuliše produkciju cAMP iz ATP. To se ostvaruje direktnom stimulacijom za membranu-vezanog enzima adenilat ciklaza. cAMP dejstvuje kao sekundarni glasnik, koji interaguje sa i aktivira proteinsku kinazu A (PKA). PKA može da fosforilizira veliki broj nizvodnih ciljeva.
- Gαi inhibira produkciju cAMP iz ATP.
- Gαq/11 stimuliše za membranu-vezanu fosfolipazu C beta, koja odseca PIP2 (mali membranski fosfoinozitol) u dva sekundarna glasnika, IP3 i diacilglicerol (DAG).
- Gα12/13 učestvuju u signalizaciji Ro familije (kroz RoGEF superfamiliju) i kontrolišu remodelovanje ćelijskog citoskeletona, i tako regulišu ćelijsku migraciju.
- Gβγ ponekad takođe ima aktivne funkcije, npr. sprezanje sa L-tipom kalcijum kanala.
Male GTPaze
Male GTPaze takođe vezuju GTP i GDP, i učestvuju u transdukciji signala. Ti proteini su homologni sa alfa (α) podjedinicom nađenom u heterotrimerima, ali postoje kao monomeri. To su mali (20-25 kDa) proteini koji vezuju guanozin trifosfat (GTP). Ova proteinska familija je homologna sa Ras GTPazama, i stoga se naziva Ras superfamilijom GTPaza.
Lipidacija
Da bi se vezali za unutrašnju stranu membrane plazme, mnogi G proteini i male GTPaze podležu lipidaciji, drugim rečima, oni su kovalentno modifikovani sa lipidnim ekstenzijama. Oni mogu biti vezani za miristinsku ili palmitinsku kiselinu, ili su prenilovani.
Reference
- ^ Reece, Jane; Campbell, Neil (2002). Biology. San Francisco: Benjamin Cummings. ISBN 0-8053-6624-5.
- ^ „G Protein Pathways -- Neves et al. 296 (5573): 1636 – Science”. 31. 5. 2002.
- ^ „The Nobel Prize in Physiology or Medicine 1994, Illustrated Lecture”.
- ^ „Press Release: The Nobel Assembly at the Karolinska Institute decided to award the Nobel Prize in Physiology or Medicine for 1994 jointly to Alfred G. Gilman and Martin Rodbell for their discovery of "G-proteins and the role of these proteins in signal transduction in cells”. 10. 10. 1994.
- ^ Wu, Ge (2010). „Assays with GPCRs”. Assay Development: Fundamentals and Practices. John Wiley and Sons. стр. 265—285. ISBN 0-470-19115-5.
- ^ Digby GJ, Lober RM, Sethi PR, Lambert NA (2006). „Some G protein heterotrimers physically dissociate in living cells.”. Proc Natl Acad Sci USA. 103 (47): 17789—94. PMC 1693825
 . PMID 17095603. doi:10.1073/pnas.0607116103.
. PMID 17095603. doi:10.1073/pnas.0607116103. - ^ Khafizov K, Lattanzi G, Carloni P (2009). „G protein inactive and active forms investigated by simulation methods”. PROTEINS : Structure, Function, and Bioinformatics. 75 (4): 919—30. PMID 19089952. doi:10.1002/prot.22303.

Literatura
- Wu, Ge (2010). „Assays with GPCRs”. Assay Development: Fundamentals and Practices. John Wiley and Sons. стр. 265—285. ISBN 0-470-19115-5.
- Reece, Jane; Campbell, Neil (2002). Biology. San Francisco: Benjamin Cummings. ISBN 0-8053-6624-5.
Dodatna literatura
- Eric R. Kandel; James H. Schwartz; Thomas M. Jessell (2000). Principles of Neural Science. New York: McGraw-Hill. ISBN 0-8385-7701-6.
- Lodish H, Berk A, Zipursky LS, Matsudaira P, Baltimore D, Darnell J (1999). Molecular Cell Biology (4th изд.). New York: Scientific American Books. ISBN 0-7167-3136-3.
- Donald Voet; Judith G. Voet (2005). Biochemistry (3 изд.). Wiley. ISBN 9780471193500.
- Bruce Alberts; Alexander Johnson; Julian Lewis; Martin Raff; Keith Roberts; Peter Walter (2002). Molecular Biology of the Cell. New York: Garlard Science. ISBN 0815332181.
- Gilman, A. G. (1987). „G proteins: transducers of receptor-generated signals”. Annu. Rev. Biochem. 56: 615—49. PMID 3113327. doi:10.1146/annurev.bi.56.070187.003151.
- Neves, S. R.; Ram, P. T.; Iyengar, Ravi (2002). „G protein pathways”. Science. 296 (5573): 1636—9. PMID 12040175. doi:10.1126/science.1071550. Текст „ Iyengar R ” игнорисан (помоћ)
- п
- р
- у
FABP1 · FABP2 · FABP3 · FABP4 · FABP5 · FABP6 · FABP7
Folistatin · Hormon rasta vezujući protein · Insulinu-sličan faktor rasta vezujući protein (IGFBP1, IGFBP2, IGFBP3, IGFBP4, IGFBP5, IGFBP6, IGFBP7) · Neurofizin (Neurofizin I, II)
Seks hormon vezujući globulin/Androgen vezujući protein · Transkortin · Tiroksin-vezujući globulin · Transtiretin
Seks hormon vezujući globulin/Androgen vezujući protein · Transkortin · Tiroksin-vezujući globulin · Transtiretin
kalcijum (Kalcijum-vezujući protein, Kalmodulin-vezujući proteini) · bakar (Ceruloplasmin) · gvožđe (Gvožđe-vezujući protein, Transferin receptor)
Retinol vezujući protein (4) · Transkobalamin
Acil prenosni protein · Adaptor protein · Holesterilester transfer protein · F-boks protein · GTP vezujući protein · Latentan TGF-beta vezujući protein · Light-žanjući kompleks · Major urinarni proteini · Membrane transport protein · Odorant vezujući protein
Нормативна контрола | |
|---|---|
| Државне |
|
| Остале |
|
G protein на сродним пројектима Википедије:








